Binocular vision deprivation study by Juhani Hyvärinen's group
In 1960s, investigations on early deprivation of vision in kittens by Torsten Wiesel and David Hubel (Nobel Price winners in 1981) showed that during the early sensitive period, deprivation leads to marked changes in the primary visual cortex: the deprived, amblyopic eye loses its usual representation in cells that should get information from both eyes. During these studies they found that if both eyes were closed for a few weeks after birth, changes in the primary visual cortex were less pronounced than when one of the eyes had been closed but the kittens were functionally blind. This finding was the starting point to the investigations of my late husband's, Juhani Hyvärinen's group on changes in brain functions after closure of eye lids of both eyes of monkeys for 7 to 11 months immediately after birth.
When the eyes were opened and the function of the cortex was investigated, visual stimulation caused strong responses in the primary visual cortex, area 17 and in area 19 along the inferior parietal gyrus. In area 19, however, less than half of the recorded cell groups responded to visual stimuli and also somatic responses were recorded, which does not occur in normal animals. These somatic responses were present when the monkey actively manipulated objects. No recording sites were activated by both visual and somatic stimuli, suggesting that the two modalities were separated from each other at this level.
In normal monkeys visual and somatic stimulation activates approximately one third of cell groups in parietal area 7. In the binocularly deprived monkeys only 1% of cell groups were activated through both modalities. Deprivation had lead to loss of representation of visual information and somatic information, especially exploration with fingers, had taken over the cell groups. This is amblyopia at a higher level than at the primary visual cortex. Activation during purely visual stimulation is present in normal animals in approximately one fourth of the cell groups. In the deprived animals it was recordable only in 2% of the cell groups and the responses were weak. The number of cell groups that responded during active somatic movements had doubled and those activated by passive somaesthetic stimulation had increased by 53% (Figure 1 A-D).
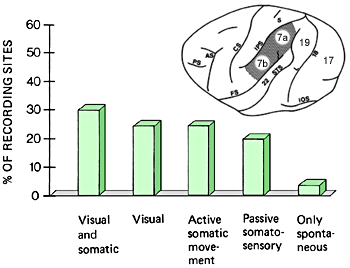
In the parietal area 7of a normal monkey several different cell groups can be found: cells that combine visual and somatic information, cell groups that are activated by only visual stimuli, those activated during active somatic movements, especially exploration by fingers or during passive somatosensory stimulation. There are very few cell groups that were not activated by any of the stimuli used in our experiments.
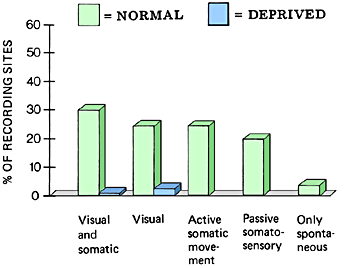
In the visually deprived monkeys the greatest changes were in the number of cell groups activated by somatic and visual information and of those activated by only visual stimuli. There were very few cell groups activated by visual or visual and somatic stimulation.
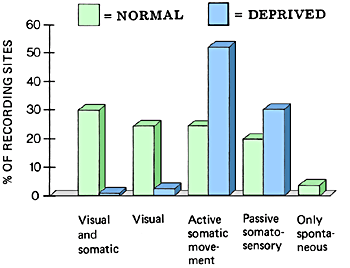
The representation of cell groups activated during active movements had increased in the visually deprived monkeys by 117% and those activated during passive somaesthetic stimulation by 53%.
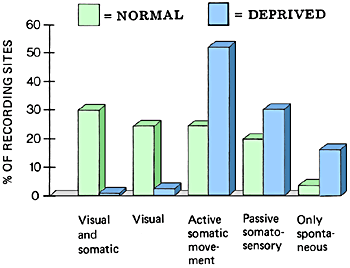
The number of cell groups that were spontaneously active but could not be activated by external stimulation and that did not change their activity during movements of the monkey increased fourfold in the visually deprived monkeys.
These findings on changes in the cortical functions of young monkeys after visual deprivation are the foundation of our present early intervention. If we want to help the visually impaired infant in an optimal way to develop visual functions, become sighted, we must start early. The associative brain functions start to develop early and are dependent on the quality of information in each of the modalities. By stressing the need to support the development of visual functions we do not forget to help the infant and child to develop all compensatory techniques. Enhancement and substitution of sight can work together.
[ Back ]