[ 1-18 ]|[ 19-37 ]
Texts of LHs lectures in Lima:
Lecture 1:
Current questions in clinical work

Slide 1. This lecture was planned to support discussion in several areas of assessment of visual functioning as classified in the ICF-CY 2007. Some routines, basic facts and recommendations ask for further discussion because they are new to the starting vision rehabilitation services. In the old services they have been a part of the work such a long time that we have forgotten how and why we started to use them in the 1980s.
When I worked as Dr. A. Edward Maumenee’s research fellow at the Wilmer Institute in late 1960s Dr. Maumenee often said to the new residents that they were very important to the development of the work of the Institute because they would ask “why some examinations or treatments were performed as they were routinely used.”
It is also today good to sometimes pause and ask the question “why”, especially because there is a change in our work in the assessment of vision for early intervention and education. With the current increase in the number of infants and children with brain damage related vision loss we should think more on the brain functions than on the eyes.
There is no “eye sight”. Sight, vision is a brain function.

Slide 2. Scientific findings seldom become a part of clinical routine examination before 20 to 25 years has past after the original report.
In our clinical assessment we are accustomed to measure with several tests WHAT the child sees, i.e. we assess visual detection and recognition functions using tests that have been developed to measure visual functions in children and adults with normal brain functions.
In early intervention, rehabilitation and special education we assess many infants and children with varying losses in brain functions and investigate HOW the child perceives and interprets his environment and tasks. The test results may depict normal brain function in a test situation or may be distorted because the needed brain function is not normal.
With the newest scientific findings, combined with earlier works we can start asking also WHY the child sees, as he seems to see. What is the nature of the vision loss?
Our questions are thus: What is the quality of the visual information entering the brain to be used for visual perception and as “vision for actions”? What can the child recognise and does he use vision for planning and executing tasks? How can we understand his experiences and learning strategies? Why can or cannot he cope with certain types of visual information?

Slide 3. When you look at these two well-known pictures you can see in the picture on the left the lower vertical surface as the side of the box closest to you BUT you can ask your brain to change the image so that the upper greyish vertical surface is closest to you. The other picture is easier to change between a vase and two profiles of faces.
Nothing changes in the incoming visual information but we can chose to see them in two different ways. This we should remember when observing and assessing children’s visual processing functions if vision is impaired and especially, if the child has brain damage related vision loss. We should never use our own vision as a measure stick.

Slide 4. The work of Torsten Wiesel and David Hubel on the structure and function of the striate cortex is well known. It started a new period in treating amblyopia and strabismus. It taught to us that a dominant function could suppress another function and prevent its development.
Torsten Wiesel compared the effect of monocular and binocular lid closure on the structure of the striate cortex and found that binocular deprivation caused less changes in the primary visual cortex than monocular closure. However, the kittens were and remained blind. This lead to the question: “If blindness is not in the primary visual cortex, were is it located.” This intrigued my late husband, Juhani Hyvärinen whose speciality was the function of the parietal lobe. It interested also me because of the great variation in the functional capacity of patients with seemingly similar disorders of eyes and/or visual pathways. We knew Torsten Wiesel from the time he was at the Karolinska Institutet in Stockholm and followed his work keenly while we were working at the Johns Hopkins University in late 1960s.

Slide 5. The location of different brain functions was mapped in many centres by carefully removing the area of the assumed functions and observing which functions had changed and which remained. In an experiment the whole occipital cortex was removed in a few monkeys. These monkeys were functionally blind in all test situations but afterwards could learn to use vision for spatial orientation. This showed that monkeys and most probably also human beings have midbrain networks with connections to visual cortical functions as shown in this slide.
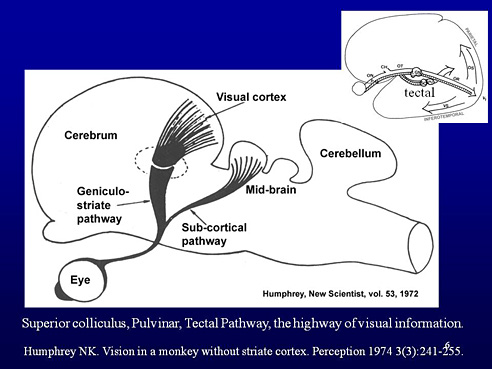
Slide 6. The subcortical visual pathway is called tectal pathway and is now known as the high-speed visual pathway that is important for ocular motor functions, especially saccades, and moving and orientation in space. Its resolving power is relatively low, i.e. small details are not perceived in motion but the larger structures in the environment are detected and recognised fast and reliably. We often forget to think on this fast pathway because we are too interested in sustained, demanding near tasks at school and occupational life.

Slide 7. My late husband and I were interested in learning more about the sensory information that we use in different tasks. His special area was tactual information in its different forms, especially important for blind people but used by everyone. My training was in the changes in vision due to retinal and pathway disorders. Back at home in Finland, my husband’s group found parietal cell groups that used information from several modalities. This was a good basis for a study on binocular deprivation of healthy newborn monkeys.

Slide 8. In this slide we see that in a normal monkey brain, the cells in the area 7a vary in their use of information (green columns). About one third of the cells use both visual and somatic information, about one fourth of the cells are only visual and another fourth are driven by active somatic movements, one fifth by passive somatosensory information and only a few cells could not be classified (it is difficult to guess what different sources of information a cell might use and therefore many different stimuli were used as long as the cell was working, caught with the microelectrode).
During visual deprivation the cells that use visual information alone or combined with information from other modalities disappeared nearly totally (blue columns).

Slide 9. The number of cells that use information from active somatic movements, especially exploration with fingers, and from passive somatosensory stimulation increased approximately as much as there was loss in the cells that were driven by vision. There was “amblyopia at a higher level”, between modalities, not between eyes. Use of vision was deprived and remained so despite the best early intervention ever arranged. The young monkeys had two caretakers and a full time psychologist for their care and activation. They developed normal motor functions with strategies typical to blind individuals.

Slide 10. This study showed that early deprivation in use of vision could lead to changes in brain functions that are permanent. Therefore, if we want to improve the use of an impaired visual system we must start early intervention really early to help the infant/child to reach the best potential functions. At the same time we help the family to develop normal, warm interactions and in the infant/ child age appropriate social development, which is the most important ability for the child’s later life. These findings are our foundation in early intervention of all developmental problems.

Slide 11. Our concept on the structure of the visual brain has changed from the Wiesel – Hubel concept to Ungerleider - Mishkin’s “What” and “Where” pathways to Milner – Goodale’s “ventral and dorsal streams”, now added with the “mirror neuron system”. These are mainly based on studies on monkey brain and on adult people who have had damage to the brain after normal development. The name “stream” does not depict the large networks where information flow is in both directions and stronger in Top-Down than toward the inferotemporal and parietal areas.
If an infant/child has had damage to the developing brain early, the development may have an unusual course due to the great plasticity of neural networks. The input of the tectal pathway is greater in the parietal lobe functions than in the temporal lobe functions although we do not yet know the connections between midbrain and primary and secondary visual cortices well enough even in the normal adult brain. There are several new techniques that are likely to enrich our understanding of function of the normal and abnormal visual brain in the future.

Slide 12. If the visual networks are drawn on the surface of the human brain, as in the above picture we see the arrows depicting the flow of visual information toward the parietal lobe via V5 and V6 as the dorsal stream (DS) that has two functionally different parts: the dorso-dorsal part is strongly motor and the dorso-ventral part combines information from several modalities for awareness of and directions in space. Visual space is represented in the activity of different cell groups, some coding location of objects in relation to the center of the visual field, some other relative to head, some third or fourth groups in relation to right or left hand. Combining the information from the different sources gives the coordinates for reaching for, grasping, etc.
Information flow for perception and recognition functions is toward the lower surface of the temporal lobe and is called ventral stream (VS). However, as stated earlier, the Top-Down flow toward the occipital lobe is stronger than the flow toward the parietal and temporal lobe. Therefore these pathways are here called networks, not streams because the flow is in two opposite directions.

Slide 13. This slide shows the original observation by di Pellegrino (di Pellegrino1992) during experiment of hand movements in a behaving monkey. Some neurons in the premotor cortex were active during goal-directed reaching of the monkey but also when the monkey saw the experimenter to perform a goal-directed reach. The cells “mirrored” the activity that they usually coded during a motor task.

Slide 14. This picture of Gallese (2007) shows the dorso-dorsal and ventro-dorsal parietal streams and the ventral stream in a monkeys brain; all three networks are connected with different areas of the frontal lobe.

Slide 15. The arrow from the anatomic area 40 to 44a depicts the third interesting network called the mirror neuron system. It was described in early 1990s and contains visual and visuomotor cell groups (Sakata et al 1995) that are active during goal-directed movements, especially those of hands and mouth but also when monkeys watch someone performing similar goal-directed movements. Findings related to mirror cell networks are made in experiments on behaving monkeys but corresponding parts of the human brain are active during movements and observation of similar movements.
Activity related to emotions can be recorded in the areas called amygdala and Insula, which is located deep in the Sylvian or lateral sulcus and is not visible to the surface of the brain.

Slide 16. The human infants are able to copy facial expressions starting at the time of birth and keenly watch facial expressions and movements of adults and other infants and children long before they can perform similar movements. By observing often-repeated activities infants learn to anticipate goal directed activities, first related to eating at the age of 5-6 months and longer activities leading to an important goal later at the age of 9 to 10 months or just before their first birthday. The brain structures that function in the human infant in the way typical to mirror neuron system of monkeys are not yet confirmed but it is obvious that these functions are important in the early communication and interaction. Infants are extremely sensitive to the emotional status of their mothers and “diagnose” depressive behavior of the mother better than medical experts. In such situations they distance themselves from the interaction turning their face away from the communication.
Damage to mirror neuron system has been suggested to be one of the causes of autism but this is difficult to prove because facial expressions cause activity in numerous areas of the brain and the goal-directed motor functions do not seem to be affected in individuals with autistic spectrum disorders.

Slide 17. A well awake newborn may copy such a complex motor task as smiling. The smile is a typical “copied expression” that we experience each day. If we smile, people around us smile back. It is a typical mirrored function. In all communication problems related to impaired vision, we can support the development of interpretation of facial expressions by bringing the infant’s hands on the face of the adult person so that the infant experiences facial expressions through tactile, haptic, kinesthetic and auditory information.

Slide 18. At the age of 12 weeks the infant and her father enjoy visual communication, which becomes an important function in the development of emotional bonding.
[ 1-18 ]|[ 19-37 ]